║├č§Ę┤Ž§╗»Š·╬█╦«╠Ä└Ēą┬ĘĮĘ©
ųąć°╬█╦«╠Ä└Ē╣ż│╠ŠW ĢrķgŻ║2016-4-10 8:41:03
╬█╦«╠Ä└Ē╝╝ąg | ģRŠ█╚½Ū“Łh▒Ż┴”┴┐Ż¼ĮĄĄ═Ų¾śIų╬╬█│╔▒Š
ĪĪĪĪ1 ę²čį(Introduction)
ĪĪĪĪ╔·╬’├ōĄ¬(Biological Nutrient RemovalŻ¼BNR)╩Ūų„┴„Ą─║═ūŅŠ▀│╔▒Šā×ä▌Ą─├ōĄ¬╝╝ągŻ¼ų„ę¬ė╔ūįBŽ§╗»║═«ÉBĘ┤Ž§╗»ā╔ļAČ╬ĮM│╔.Ę┤Ž§╗»▀^│╠┐╔ęįį┌╚▒č§║═ģÆ觌l╝■Ž┬▀MąąŻ¼ü厧╦ß¹}Ą¬Ž╚▒╗▀ĆįŁ×ķŽ§╦ß¹}Ą¬Ż¼Ž§╦ß¹}Ą¬į┘▒╗▀ĆįŁ×ķÜŌæBĄ¬Ż¼ūŅĮKīŹ¼FĄ¬Ą─╚ź│²Ż¼Ą½č§┐╔ęį╚Ī┤·ü厧╦ß¹}║═Ž§╦ß¹}ū„×ķļŖūė╩▄¾w╩╣Ę┤Ž§╗»▀^│╠╩¦ą¦.ę“×ķé„Įy├ōĄ¬└ĒšōšJ×ķŻ¼č§ū„×ķļŖūė╩▄¾w╩Ūā׎╚ė┌ü厧╦ß¹}║═Ž§╦ß¹}Ą─Ż¼║¾š▀▓╗į┘╩Ū─®Č╦ļŖūė╩▄¾wŻ¼▀MČ°╩╣Ę┤Ž§╗»▀^│╠▒╗ęųųŲ.ļSų°╔·╬’├ōĄ¬└Ēšō蹊┐Ą─▀Mę╗▓Į╔Ņ╚ļŻ¼20╩└╝o80─Ļ┤·Ż¼Thiosphaera pantotrophaū„×ķĄ┌ę╗ĘN║├č§Ę┤Ž§╗»Š·į┌├ō┴“Ę┤Ž§╗»ŽĄĮyųą▒╗░l¼FŻ¼▓óūC├„┤µį┌║├č§Ę┤Ž§╗»├ĖŽĄĮy.į┌▀@ę╗蹊┐│╔╣¹Ą─╗∙ĄA╔ŽŻ¼ą┬Ą─║├č§Ę┤Ž§╗»Š·▓╗öÓ▒╗░l¼F║═Ęųļx│÷üĒ.Č°║├č§Ę┤Ž§╗»└Ēšō┐╔ęį║å╩÷×ķŻ¼į┌║├č§Ę┤Ž§╗»▀^│╠ųąŻ¼ėąÖC╠╝į┤×ķļŖūė╣®¾wŻ¼č§Īóü厧╦ß¹}║═Ž§╦ß¹}Š∙×ķļŖūė╩▄¾wŻ¼ūŅĮKŽ§╦ß¹}ŅÉ▒╗▐D╗»×ķÜŌæBĄ¬;▀@ę╗▀^│╠ųąŲū„ė├Ą─├Ė×ķŽ§╦ß¹}▀ĆįŁ├ĖĪóü厧╦ß¹}▀ĆįŁ├ĖĪóę╗č§╗»Ą¬▀ĆįŁ├Ė║═ę╗č§╗»Č■Ą¬▀ĆįŁ├Ė.ę“┤╦║├č§Ę┤Ž§╗»Ą─╠ž³c┐╔ęįÜw╝{×ķŻ║ó┘Ę┤Ž§╗»▀^│╠į┌║├觌l╝■Ž┬▀Mąą┐╔īŹ¼F═¼▓ĮŽ§╗»Ę┤Ž§╗»;ó┌Ž§╗»«a╬’┐╔ęįų▒Įėū„×ķĘ┤Ž§╗»Ąū╬’▒╗└¹ė├ęį▒▄├ŌŽ§╦ß¹}ŅÉ╬’┘|Ą─Ęe└█;ó█Ę┤Ž§╗»╔·│╔Ą─OH-┐╔ęį▓┐Ęųča│õŽ§╗»ū„ė├Ž¹║─Ą─ēAČ╚ęįŠS│ųpHĄ─ŽÓī”ĘĆČ©.
ĪĪĪĪĮ³─ĻüĒŻ¼ī”║├č§Ę┤Ž§╗»Ą─蹊┐ų„ę¬╝»ųąį┌╠žČ©║├č§Ę┤Ž§╗»Š·ųĻĄ─Ęųļx║═ŲõĘ┤Ž§╗»╠žąįĄ─蹊┐Ż¼Ą½╠žČ©Š·ųĻĄ─Ę┤Ž§╗»╠žąį┼c╠╝į┤ŅÉą═Īó╠╝Ą¬▒╚Īó£žČ╚Īó╚▄ĮŌč§║═pHĄ╚ę“╦žėąĻP;Č°Ūę▓╗═¼Ą¬į┤ę▓ė░Ē抷ųĻĄ─Ę┤Ž§╗»╠žąį.Å─┐╔─▄Ą─╣ż│╠īŹļHæ¬ė├ĮŪČ╚┐╝æ]Ż¼ī”║├č§Ę┤Ž§╗»Š·╚║š¹¾w║═ŲõĘ┤Ž§╗»╠žš„æ¬▀Mę╗▓Į蹊┐╩Ūėą▒žę¬Ą─Ż¼▒Ššō╬─Ą─蹊┐─┐Ą─╝┤į┌ė┌┤╦.į┌Ž╚Ū░Ą─▀B└m┴„A/O┴„ļxĘ┤æ¬Ų„═¼▓ĮŽ§╗»Ę┤Ž§╗»čąŠ┐▀^│╠ųą░l¼FŻ¼║├č§Ę┤Ž§╗»▀^│╠į┌═¼▓ĮŽ§╗»Ę┤Ž§╗»ųąŲĄĮĘŪ│Żųžę¬Ą─ū„ė├Ż¼čąŠ┐ĮY╣¹ę▓ūC├„┴╦║├č§Ę┤Ž§╗»Š·Ą─┤µį┌.ę“┤╦ėą▒žę¬ęįš¹¾wŠ·╚║×ķ蹊┐ī”Ž¾Ż¼į┌╠žČ©╦«ŁhŠ│Śl╝■Ž┬ī”▓╗═¼Ą¬į┤ī”╣”─▄╝ÜŠ·Ą─╗Ņąįė░Ēæ║═Ę┤Ž§╗»╠žš„▀MąąčąŠ┐Ż¼╩╣║├č§Ę┤Ž§╗»Š·║═║├č§Ę┤Ž§╗»ū„ė├į┌╬█╦«ų╬└ĒųąŠ▀ėąīŹļHæ¬ė├Ą─┐╔─▄ąį.
ĪĪĪĪ2 ▓─┴Ž┼cĘĮĘ©
ĪĪĪĪ2.1 įć“×čbų├
ĪĪĪĪłD 1×ķ▓╔ė├┴„ļx╔·╬’╝╝ągĄ─ę╗╠ūąĪą═┴„ļx╔·╬’─żĘ┤æ¬Ų„.Ę┤æ¬Ų„×ķå╬Ė±╩ęŻ¼ėąą¦╚▌Ęe3.2 LŻ¼Ė±╩ęĮYśŗ┼c╬─½I19Ą─A/O┴„ļxĘ┤æ¬Ų„å╬ę╗Ė±╩ęŅÉ╦ŲŻ¼╠Ņ┴Ž╩Ū╚ĪūįA/OĘ┤æ¬Ų„║├č§ģ^ā╚Ą─┴„ļxŪ“Ż¼┴„ļxŪ“ų▒ÅĮ.Ę┤æ¬Ų„Ž┬▓┐│÷╦«Ż¼╔Ž▓┐▀M╦«Ż¼╚õäė▒├┐žųŲ.▓╔ė├Ąū▓┐ŲžÜŌ▒PŲžÜŌŻ¼ŲžÜŌ┴┐┐žųŲ║═ėŗ┴┐▓╔ė├╬óļŖ─XŲžÜŌčbų├┐žųŲŻ¼═©▀^╠ĮŅ^Š½┤_£y┴┐║═╬óš{Ż¼īŹ¼Fįć“×╦∙ę¬Ū¾Ą─Ė╗č§╦«ŁhŠ│Śl╝■.£žČ╚┐žųŲ▓╔ė├╝ė¤ß░¶īŹ¼F.
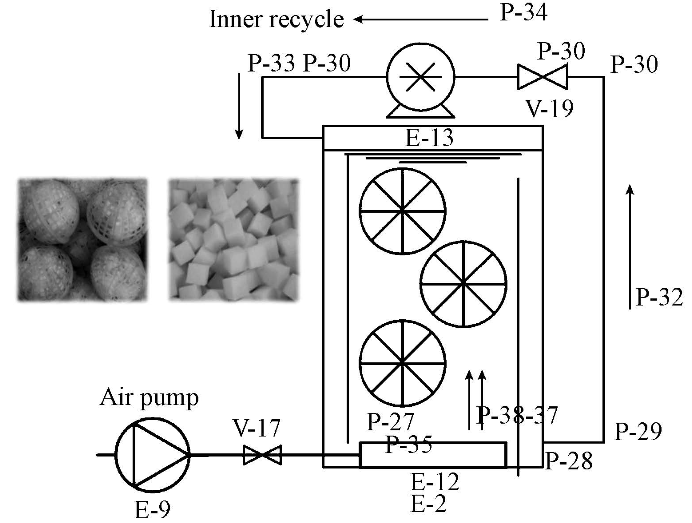
ĪĪĪĪłD1 ąĪą═┴„ļx╔·╬’─żĘ┤æ¬Ų„╩ŠęŌłD
ĪĪĪĪ┴„ļx¼FŽ¾Ą─ĮŌßīŻ║┴„¾wį┌┴„äėųą┐é┤µį┌ų°▓╗═¼Ą─┴„╦┘┐ņ║═┴„╦┘┬²Ą─ł÷╦∙Ż¼╣╠¾w╬’║═ėąÖC╬’─z¾wį┌┴„¾wĄ─┴„äėųąŻ¼┐é╩Ūė╔┴„╦┘┐ņĄ─ę╗é╚Ž“┴„╦┘┬²Ą─ę╗é╚╝»ųąŠ█╝»Ż¼▀@ĘN¼FŽ¾ĘQų«×ķĪ░┴„ļxĪ▒.┴„ļx╔·╗»╠Ä└Ē╝╝ąg╝┤╩Ūį┌¤oē║┴”Īóų╗ąĶ╦«¾w╔į╬ó┴„äėĪó╠Ņ┴Ž×ķ▒Ē├µĮø▀^╠ž╩Ō╠Ä└ĒĄ─▓─┴ŽĄ─╝»║Ž¾w(┴„ļxŪ“)ųąīŹ¼F.╬█╦«į┌┴„äėųą┤µį┌ų°Ū“¾w═Ō┴„╦┘┐ņŻ¼Ū“¾wā╚┴„╦┘┬²Ą─ĀŅørŻ¼╬█╦«ųąŲ»ĖĪ╬’╝»ųąį┌┴„╦┘┬²Ą─ĄžĘĮ«a╔·┴„ļx¼FŽ¾.Įø▀^¤oöĄ┤╬┴„ļxū„ė├Ż¼╩╣╬█╦«ųąĄ─╣╠¾w╬’║═ėąÖC╬’─z¾w┼c╦«ĘųļxŻ¼į┘ĮY║Ž╔·╗»ĘųĮŌŻ¼śŗ│╔┴╦┴„ļx╔·╗»╝╝ąg.įć“×ė├Ę┤æ¬Ų„╝┤ę└ššįōįŁ└ĒśŗĮ©Īó▀\ąą.
ĪĪĪĪįć“×ė├╦«Ęųäe×ķĄ═╠╝Ą¬▒╚ÅU╦«(ęį░▒Ą¬×ķĄ¬į┤Ą─ÅU╦«)ĪóŽ§╦ß¹}ÅU╦«║═ü厧╦ß¹}ÅU╦«.Ą═╠╝Ą¬▒╚ÅU╦«▓╔ė├ūįüĒ╦«═Č╝ė┤ū╦ßŌcĪó┬╚╗»õ@Īó┴ū╦ßČ■ÜõŌø┼õųŲ;Ž§╦ß¹}║═ü厧╦ß¹}┼õ╦«▓╔ė├ūįüĒ╦«═Č╝ė┬╚╗»õ@ĪóŽ§╦ßŌøĪóü厧╦ßŌc┼õųŲ─ŻöMĖ▀╠╝Ą¬▒╚ÅU╦«Ż¼╦Ää®═Č╝ė┴┐ęįØMūŃįć“×ę¬Ū¾Ą─ØŌČ╚ĘČć·×ķ£╩.ĘųŅÉ╦«┘|║═┐žųŲ£žČ╚╚ń▒Ē 1╦∙╩Š.
ĪĪĪĪ▒Ē1 įć“×įŁ╦«╦«┘|║═£žČ╚Śl╝■
ĪĪĪĪ2.2 │ŻęÄĘų╬÷ĘĮĘ©
ĪĪĪĪ╦«śė£yČ©Ū░Įø0.45 ”╠m×V╝ł▀^×VŻ¼NH4+-NĪóNO2--NĪóNO3--Nę└ō■ś╦£╩ĘĮĘ©Öz£y(ć°╝ęŁhŠ│▒Żūo┐éŠųŻ¼2002)Ż¼TN║═TCĪóTOCĪóTIC╩╣ė├vario TOCįOéõ(ElementarŻ¼Ą┬ć°)£yČ©.DOĪópHĪó£žČ╚▓╔ė├į┌ŠĆ╠ĮŅ^▒O£y(WTWŻ¼Ą┬ć°).
ĪĪĪĪ2.3 16S rDNA╠ß╚ĪĪóPCRöUį÷Īó┐╦┬Ī£yą“╝░ŽĄĮy░lė²Ęų╬÷
ĪĪĪĪDNA╠ß╚Ī▓╔ė├╔Ž║Ż╔·╣ż╔·«aĄ─įćä®║ąŻ¼PCRöUį÷╦∙ė├ę²╬’╚ń▒Ē 2╦∙╩Š.
ĪĪĪĪ▒Ē2 PCRöUį÷╠Įßś
ĪĪĪĪĘ┤Ž§╗»Š·╩╣ė├ę²╬’nirS1F║═nirS6Rī”Ę┤Ž§╗»Š·▀MąąöUį÷.PCRĘ┤款wŽĄ(50 ”╠L)×ķŻ║5 ”╠LĄ─10Ī┴PCR bufferŻ¼dNTP(Ė„2.5 mmol Īż L-1)2 ”╠LŻ¼nirS1F(20 ”╠mol Īż L-1)1 ”╠LŻ¼nirS6R(20 ”╠mol Īż L-1)1 ”╠LŻ¼Taq├Ė(5U)1 ”╠LŻ¼DNA─Ż░Õ2 ”╠LŻ¼╝ėddH2Oų┴50 ”╠L.PCR▓╔ė├ĮĄ┬õ╩ĮöUį÷│╠ą“Ż¼Š▀¾wĘ┤æ¬Śl╝■×ķŻ║95 ĪµŅAūāąį5 min;95 Īµūāąį30 sŻ¼60 Īµ═╦╗45 sŻ¼72 Īµčė╔ņ1 minŻ¼╣▓▀Mąą20éĆ裣hŻ¼├┐éĆ裣hĮĄĄ═0.5 Īµ;95 Īµūāąį30 sŻ¼50 Īµ═╦╗45 sŻ¼72 Īµčė╔ņ1 minŻ¼╣▓▀Mąą10éĆ裣h;72 Īµčė╔ņ10 min.
ĪĪĪĪī”PCRöUį÷«a╬’▀MąąŪą─z╝ā╗»Ż¼īóPCR╗ž╩š«a╬’┼cpMD18-T▌d¾w▀Mąą▀BĮė║¾Ż¼▐D╚ļJM109Ėą╩▄æB╝Ü░¹Ż¼ūŅ║¾▀Mąą╦{░ū░▀║Y▀x.╠¶╚ĪĻ¢ąį┐╦┬ĪūėŻ¼╦══∙╔Ž║Ż╔·╣ż▀Mąą£yą“.īó╦∙Ą├ą“┴ą└¹ė├BLAST│╠ą“┼cGenBankųąęčĄŪõøĄ─ą“┴ą▀Mąą═¼į┤ąį▒╚▌^Ż¼▓ó└¹ė├MEGA4.0▄ø╝■ųąĄ─ÓÅĮė╦ŃĘ©śŗĮ©ŽĄĮy░lė²śõ.
ĪĪĪĪ2.4 ¤╔╣ŌįŁ╬╗ļsĮ╗╝╝ąg(FISH)
ĪĪĪĪFISH╝╝ągė├ė┌Ęų╬÷╗Ņąį╝ÜŠ·Ą─ŽÓī”öĄ┴┐ęį▀_ĄĮ┼ąöÓŽĄĮy▀\ąąĀŅørĄ──┐ś╦.░▒č§╗»Š·▓╔ė├NSO1225(CGCCATTGTATTACGTGTGA)╠ĮßśĪóü厧╗»Š·▓╔ė├NIT3(CCTGTGCTCCATGCTCCG)╠ĮßśÖz£y(Schmid et al., 2000).ļsĮ╗▓Į¾E╚ńŽ┬Ż║
ĪĪĪĪ(1)╬█─Ó╚ĪśėŻ║┴„ļx╠Ņ┴ŽĮø│¼┬Ģ║¾īóäāļxĄ─╬█─Ó╚Ī2Ī½5 mLų┴ļxą─╣▄Ż¼2000 r Īż min-1ļxą─5 minŻ¼╚ź╔ŽŪÕę║(╝ėš¶s╦«ųžÅ═ā╔┤╬);śėŲĘ╝ė╚ļ1 mLČÓŠ█╝ū╚®▓óōuä“Ż¼4 ĪµŽ┬Ę┼ų├3 h.śėŲĘ12000 r Īż min-1ļxą─5 minŻ¼╚ź╔ŽŪÕę╣;╝ė╚ļ1Ī┴PBSōuä“Ż¼10000 r Īż min-1ļxą─5 minŻ¼╚ź╔ŽŪÕę╣(ųžÅ═3┤╬);╝ė0.5 mLĄ─1Ī┴PBSĪó0.5 mL¤o╦«ęę┤╝Ż¼ōuä“║¾-20 Īµ▒Ż┤µ.
ĪĪĪĪ(2)śėŲĘ╣╠Č©Ż║ŽĪßīśėŲĘ3~5▒ČŻ¼ī”śėŲĘ▀Mąą│¼┬Ģ╠Ä└ĒŻ¼īó╬█─Óą§¾w┤“╔ó│╔å╬éĆ╝Ü░¹ęį▒Ńė┌’@╬óńRėŗöĄ.╚╗║¾╚Ī3 ”╠LśėŲĘ═┐ė┌░³┬±├„─zĄ─▓ŻŲ¼╔Ž(Öz▓ķśėŲ¼▒ŠĄū)Ż¼37 ĪµĄ─¤ß║µŽõ╣╠Č©2 h.ę└┤╬ė├┘|┴┐▒╚50%Īó80%Īó98%Ą─ęę┤╝Į■Øn3 minŻ¼ī”╝Ü░¹▀Mąą├ō╦«▓óĖ╔į’.
ĪĪĪĪ(3)śėŲĘļsĮ╗Ż║╬³╚Ī2 mLļsĮ╗ŠÅø_ę║▒ķ▓╝į┌ļsĮ╗║ąā╚š█║├Ą─╬³╦«╝ł╔ŽŻ¼īóęč╣╠Č©║├śėŲĘĄ─▌d▓ŻŲ¼Ę┼╚ļļsĮ╗╣▄ųąŻ¼╚╗║¾į┌46 ĪµļsĮ╗ĀtųąĘ┼ų├öĄmin;╬³╚Ī10 ”╠L╠Įßś┘A┤µę║║═80 ”╠LļsĮ╗ŠÅø_ę║(Hybridization BufferŻ¼HB║═Washing BufferŻ¼WB)Ż¼╗ņ║Ž║¾ė├▓Ł╝ł░³║├Ę┼╚ļ46 ĪµļsĮ╗ĀtųąŅA¤ßöĄmin;╠Įßś┘A┤µę║ØŌČ╚×ķ25 ng Īż ”╠L-1Ż¼ė├¤oŠ·╦«ŽĪßī┘Å┘IĄ─╠ĮßśŻ¼╬³╚Ī9 ”╠LŅA¤ß║¾Ą─╠ĮßśŽĪßīę║═┐ė┌▌d▓ŻŲ¼┤²£yśėŲĘ╔ŽŻ¼╚╗║¾īó▌d▓ŻŲ¼čĖ╦┘ĄžęŲ╗žļsĮ╗╣▄ųąė┌46 ĪµŽ┬▀MąąļsĮ╗2~3 h;ļsĮ╗║¾┤“ķ_║Ń£ž╦«įĪ▓█Ż¼╝ė¤ßĄĮ48 ĪµŻ¼ī”ļsĮ╗ŠÅø_ę║Īó┴▄Ž┤ŠÅø_ę║▀MąąŅA¤ß.ļsĮ╗║ąųą╚Ī│÷▌d▓ŻŲ¼ė├ļsĮ╗ŠÅø_ę║ø_Ž┤śėŲĘ║¾Ż¼┐ņ╦┘Ę┼╚ļ┴▄Ž┤ŠÅø_ę║Ż¼48 Īµ╦«įĪ20 min║¾ė├4 Īµ▒∙╦«ø_Ž┤śėŲĘŻ¼śėŲĘØŹā¶┼_ųąō]Ė╔.
ĪĪĪĪ(4)ĘŌŲ¼ė^▓ņŻ║═┐ĘŌŲ¼ä®Ż¼╔w╔w▓ŻŲ¼Ż¼¤oÜŌ┼▌║¾ųĖ╝ūė═ĘŌčb;ė├Ħėą360 nm╝ż░l▓©ķLĪó460 nm░l╔õ▓©ķLĄ─×V╣ŌŲ¼Ą─¤╔╣Ō’@╬óńRė^▓ņ╝Ü░¹.
ĪĪĪĪ3 įć“×ĮY╣¹┼cėæšō
ĪĪĪĪ3.1 Ą═╠╝Ą¬▒╚ÅU╦«║├č§Ę┤Ž§╗»čąŠ┐
ĪĪĪĪ3.1.1 Ę┤æ¬Ų„▀\ąąĀŅæB
ĪĪĪĪłD 2a╦∙╩Š×ķĘ┤æ¬Ų„ā╚│ų└m24 hĄ─DO║═pHūā╗»┌ģä▌.ė╔ė┌║├č§Ę┤Ž§╗»Š·ī”DOØŌČ╚▓╗├¶ĖąŻ¼ŠS│ų▌^Ė▀DOØŌČ╚▓ó▓╗ė░Ēæ║├č§Ę┤Ž§╗»Š·Ą─╔·ķL║═╗Ņąį.═¼ĢrpH▓ó╬┤│÷¼F┤¾Ą─▓©äėŻ¼į┌š¹éĆįć“×▀^│╠ųą╩╝ĮKŠS│ųį┌7.0~8.0ų«ķgŻ¼▀@ę╗pHĘČć·▀mę╦Ž§╗»Š·║═Ę┤Ž§╗»Š·Ą─╔·┤µŻ¼Ų½ēAąįŁhŠ│▒Ē├„┤µį┌▌^║├Ą─Ę┤Ž§╗»ū„ė├.£žČ╚ĘĆČ©┐žųŲį┌25~30 Īµ.
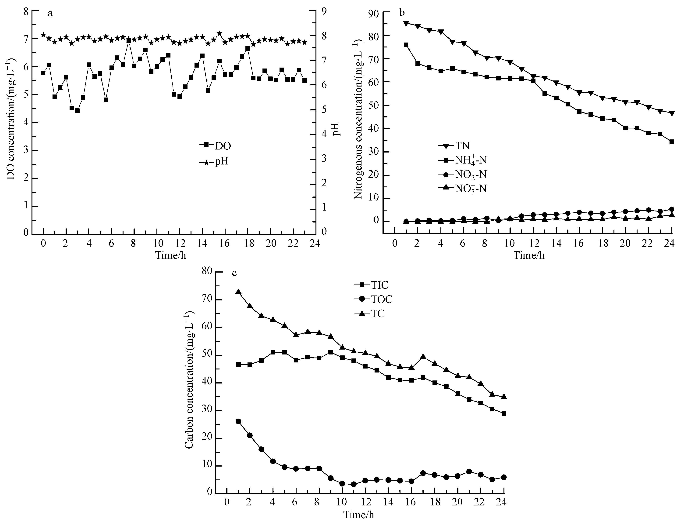
ĪĪĪĪłD2 ▀\ąąģóöĄ║═╬’┘|ØŌČ╚ūā╗»┌ģä▌
ĪĪĪĪ3.1.2 ╬’┘|╚ź│²▀^│╠Ęų╬÷
ĪĪĪĪĄ═╠╝Ą¬▒╚ÅU╦«┐╔ęį┐┤ū„ęį░▒Ą¬×ķų„ꬥ¬į┤Ą─ÅU╦«.łD 2b║═2cĘųäe×ķĄ¬ŅÉ╬’┘|║═╠╝ŅÉ╬’┘|╚ź│²╝░▐D╗»┌ģä▌.ė╔łD 2b┐╔ų¬Ż¼┐饬║═░▒Ą¬ØŌČ╚čžĘ┤æ¬ĢrķgųØuŽ┬ĮĄŻ¼Ū·ŠĆą▒┬╩┤¾ė┌│Ū╩ą╬█╦«įć“×;ü厧╦ß¹}║═Ž§╦ß¹}į┌Ę┤æ¬▀Mąą10 h║¾│÷¼F╔┘┴┐Ęe└█Ż¼Ž§╦ß¹}Ęe└█┴┐ąĪė┌ü厧╦ß¹}.ī”æ¬łD 2cŻ¼ėąÖC╠╝ØŌČ╚į┌0~10 hų«ķg┐ņ╦┘ĮĄĄ═Ż¼Č°¤oÖC╠╝ūā╗»Ę∙Č╚ŽÓī”║▄ąĪ.▀@ę╗┌ģä▌▒Ē├„Ż¼0~10 hā╚Ą─░▒Ą¬╚ź│²┐╔─▄üĒūį«ÉBŽ§╗»ū„ė├Ż¼║├č§Ę┤Ž§╗»ū„ė├īóŽ§╗»ū„ė├╔·│╔Ą─Ž§╦ß¹}ŅÉ╬’┘|╚ź│².įŁę“╚ńŽ┬Ż║ę╗╩Ūė╔ė┌▓┐Ęų║├č§Ę┤Ž§╗»Š·ę▓╩Ū«ÉBŽ§╗»Š·Ż¼į┌Ą═╠╝Ą¬▒╚╬█╦«├ōĄ¬▀^│╠ųąŻ¼«ÉBŽ§╗»Š·┼cūįBŽ§╗»Š·ĖéĀÄĄ¬į┤Ż¼░▒Ą¬Ą─╚ź│²ų„ę¬ė╔«ÉBŽ§╗»Š·═Ļ│╔Ż¼ę“┤╦ėąÖC╠╝▒╗┤¾┴┐Ž¹║─Ż¼Č°¤oÖC╠╝ØŌČ╚Ž┬ĮĄ┌ģä▌ąĪė┌ėąÖC╠╝.Č■╩Ū┐╔─▄═¼Ģr┤µį┌ü厧╦ß¹}▀ĆįŁ├Ė║═Ž§╦ß¹}▀ĆįŁ├ĖŻ¼Ž§╗»▀^│╠«a╔·Ą─Ž§╦ß¹}ŅÉ╬’┘|į┌Ė▀╗Ņąįü厧╦ß¹}▀ĆįŁ├Ė║═Ž§╦ß¹}▀ĆįŁ├Ėū„ė├Ž┬▒╗čĖ╦┘▀ĆįŁ.įć“×ųą▀Ć░l¼FŻ¼1~2 hĄ─░▒Ą¬ėą═╗ĮĄŻ¼┐╔─▄╩Ū┤µį┌─│ĘNūįBŽ§╗»Š·╩╣╔┘┴┐░▒Ą¬į┌įć“×│§Ų┌▒╗ų▒Įė▐D╗»×ķÜŌæBĄ¬╚ź│²Ż¼Ą½Ģ║Ģr╚▒Ę”įć“×ų¦│ųŻ¼ąĶ▀Mę╗▓Į蹊┐.┐éų«Ż¼0~10 hĘ┤æ¬Ų„ā╚│÷¼F┴╦═¼▓ĮŽ§╗»Ę┤Ž§╗»¼FŽ¾Ż¼Č°Ūęų„ę¬┐╔─▄üĒūį«ÉBŽ§╗»-║├č§Ę┤Ž§╗»ū„ė├.
ĪĪĪĪį┌10~24 hļAČ╬Ż¼łD 2c’@╩ŠėąÖC╠╝Ž┬ĮĄ┌ģä▌╗∙▒ŠĮY╩°Ż¼Ą½╩Ū¤oÖC╠╝ØŌČ╚ķ_╩╝Ž┬ĮĄ.ī”æ¬łD 2b╦∙╩ŠŻ¼10~24 h░▒Ą¬ØŌČ╚└^└mŽ┬ĮĄŻ¼Ž§╦ß¹}║═ü厧╦ß¹}ķ_╩╝«a╔·ąĪĘ∙Č╚Ą─Ęe└█.Ęų╬÷įŁę“Ż║ę╗╩Ūė╔ė┌ėąÖC╠╝▒╗┤¾┴┐Ž¹║─║¾Ż¼«ÉBŽ§╗»Š·Ą─į÷ų│╩▄ĄĮ╠╝į┤▓╗ūŃĄ─ė░ĒæŻ¼╗ŅąįĮĄĄ═;Č°ūįBŽ§╗»Š·į÷ų│╦┘Č╚╝ė┐ņŻ¼╗Ņąįį÷ÅŖŻ¼ī”¤oÖC╠╝į┤Ą─Ž¹║─į÷┤¾Ż¼▒Ē├„▀@ę╗ļAČ╬Ą─░▒Ą¬╚ź│²ų„ę¬üĒūįūįBŽ§╗»ū„ė├.Č■╩Ūė╔ė┌ėąÖC╠╝į┤Ą─▓╗ūŃ╩╣║├č§Ę┤Ž§╗»Š·ī”Ž§╦ß¹}ŅÉ╬’┘|Ą─╚ź│²─▄┴”ę▓▒╗Ž„╚§;Ą½╩Ūė╔ė┌ūįBŽ§╗»Š·┐╔ęį└¹ė├NH3č§╗»Ą──▄┴┐▀ĆįŁCO2×ķėąÖC╠╝╣®╔·├³╗ŅäėŻ¼ę╗Č©│╠Č╚╔Žę▓ča│õ┴╦ėąÖC╠╝Ą─Ž¹║─Ż¼║├č§Ę┤Ž§╗»Š·┐╔ęį└^└m▒Ż│ųę╗Č©Ą─Ę┤Ž§╗»ū„ė├Ż¼Ą½Ę┤Ž§╗»│╠Č╚╩▄Ž▐Ż¼ę“┤╦┐╔─▄ī¦ų┬Ž§╦ß¹}ŅÉ╬’┘|Ą─ąĪĘ∙Ęe└█¼FŽ¾.╚²╩Ūü厧╦ß¹}╩ŪŽ§╦ß¹}Ę┤Ž§╗»▀^│╠Ą─ųąķg«a╬’Ż¼ŲõØŌČ╚▀^Ė▀ęūī”Ž§╦ß¹}▀ĆįŁ×ķü厧╦ß¹}▀@ę╗▓Į¾E«a╔·ęųųŲū„ė├Ż¼Å─Č°ĮĄĄ═Ž§╦ß¹}Ą─▀ĆįŁ┴┐Ż¼Ą½ü厧╦ß¹}ØŌČ╚▀_ĄĮČÓ╔┘╝┤ė░Ē控╦ß¹}Ą─▀ĆįŁŻ¼─┐Ū░Ģ║Ģr¤oöĄō■ų¦ō╬.Č°Ž§╦ß¹}Ą─╔┘┴┐Ęe└█Ż¼▒Ē├„ü厧╦ß¹}Ęe└█Ą─ØŌČ╚ęčĮø▀_ĄĮė░Ēæ║├č§Ę┤Ž§╗»Š·ī”Ž§╦ß¹}Ę┤Ž§╗»─▄┴”Ą─│╠Č╚.
ĪĪĪĪį┌▀B└m24 hĄ─Ą═╠╝Ą¬▒╚║├č§Ę┤Ž§╗»čąŠ┐ųąŻ¼Ę┤æ¬ļAČ╬▒╗Ęų×ķ«ÉBŽ§╗»-║├č§Ę┤Ž§╗»║═ūįBŽ§╗»-║├č§Ę┤Ž§╗»ā╔ļAČ╬Ż¼Ą½Š∙Ę¹║Ž═¼▓ĮŽ§╗»Ę┤Ž§╗»╠žš„.Ė∙ō■ėŗ╦Ń┐╔ų¬Ż¼0~10 hĄ─═¼▓ĮŽ§╗»Ę┤Ž§╗»┬╩×ķ79.40%Ż¼10~24 hĄ─═¼▓ĮŽ§╗»Ę┤Ž§╗»┬╩×ķ81.25%.
ĪĪĪĪ3.1.3 ¤╔╣ŌįŁ╬╗ļsĮ╗įć“×
ĪĪĪĪęį╔ŽĄ─║├č§Ę┤Ž§╗»╠žš„Ęų╬÷ęį╝░╔┘┴┐Ą─Ž§╦ß¹}ŅÉ╬’┘|Ęe└█¼FŽ¾ę└┼f┤µį┌ę╗éĆå¢Ņ}Ż¼Š═╩Ūė^▓ņĄĮĄ─ü厧╦ß¹}Ęe└█╩Ūʱę╗Č©üĒūį║├č§Ę┤Ž§╗»─▄┴”▒╗Ž„╚§▓┼«a╔·Ą─Ż¼▀Ć╩Ūė╔ūįBŽ§╗»ū„ė├╔·│╔Ż¼▀@ąĶę¬═©▀^Ųõ╦³čąŠ┐╩ųČ╬üĒ┼ąöÓ.łD 3×ķĄ┌23 hĄ─░▒č§╗»Š·(AOB)║═ü厧╗»Š·(NOB)Ą─ŽÓī”öĄ┴┐ī”▒╚Ż¼Ę┼┤¾▒ČöĄ×ķ120▒Č.Å─ŠG╔½╣ŌļŖöĄ┴┐üĒ┐┤Ż¼Č■š▀╗ŅąįŠ·öĄ┴┐╗∙▒ŠĮėĮ³Ż¼šf├„ü厧╦ß¹}│÷¼FĘe└█▓ó▓╗╩ŪüĒūįūįBŽ§╗»ū„ė├Ż¼Č°╩Ū║├č§Ę┤Ž§╗»Š·Ę┤Ž§╗»─▄┴”╩▄Ž▐Ż¼ü厧╦ß▀ĆįŁ├Ė╗Ņąį▒╗ęųųŲ▀MČ°ė░Ēæ┴╦ü厧╦ߥ─▀ĆįŁŻ¼ę“┤╦│÷¼FąĪĘ∙Ęe└█¼FŽ¾.

ĪĪĪĪłD3 AOB║═NOBŽÓī”öĄ┴┐▒╚▌^
ĪĪĪĪ3.2 ęįŽ§╦ß¹}×ķĄ¬į┤ÅU╦«Ą─║├č§Ę┤Ž§╗»čąŠ┐
ĪĪĪĪ3.2.1 ╬’┘|╚ź│²▀^│╠┼cĘų╬÷
ĪĪĪĪłD 4a║═4bĘųäe×ķĄ¬ŅÉ╬’┘|║═╠╝ŅÉ╬’┘|╚ź│²╝░▐D╗»┌ģä▌.ė╔łD 4a┐╔ų¬Ż¼į┌0~6 hļAČ╬Ż¼Ž§╦ß¹}ŠÅ┬²Ž┬ĮĄČ°ü厧╦ß¹}į┌ķ_╩╝ąĪĘ∙╔Ž╔²║¾╝┤╗∙▒Š▓╗ūā.▀@ę╗ļAČ╬ūā╗»▒Ē├„Ż¼ė╔ė┌┴„ļx╠Ņ┴Ž╚Īūį│Ū╩ą╬█╦«Ę┤æ¬Ų„Ż¼įŁ╦«Ą─Ą¬į┤╠žš„░l╔·Ė─ūāŻ¼║├č§Ę┤Ž§╗»Š·ąĶ꬚{š¹ęį▀mæ¬ą┬Ą─Ą¬į┤ŁhŠ│;Č°ü厧╦ß¹}╬┤«a╔·Ęe└█┐╔─▄╩Ūü厧╦ß¹}▀ĆįŁ├Ėį┌Ž§╦ß¹}▀ĆįŁ├Ė═¼Ģr┤µį┌ĢrŠ▀ėą▌^Ė▀Ą─Ę┤æ¬╗ŅąįŻ¼ĮøŽ§╦ß¹}▀ĆįŁ├Ė«a╔·Ą─ü厧╦ß¹}į┌▌^Ė▀╗ŅąįĄ─ü厧╦ß¹}▀ĆįŁ├Ėū„ė├Ž┬▒╗čĖ╦┘▀ĆįŁ;═¼Ģr▀@ę╗ļAČ╬ėąÖC╠╝Ą─Ž¹║─┴┐ę▓▌^Ą═(łD 4b).
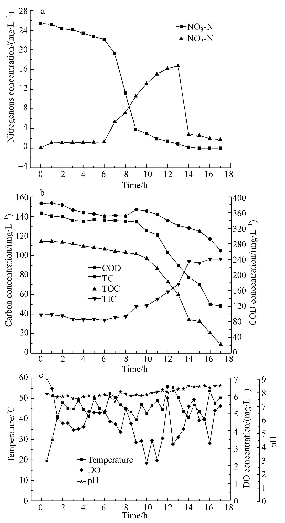
ĪĪĪĪłD4 ╬’┘|ØŌČ╚║═▀\ąąģóöĄūā╗»┌ģä▌
ĪĪĪĪłD 4a╦∙╩ŠŻ¼į┌6~9 hļAČ╬Ż¼Ž§╦ß¹}▒╗čĖ╦┘╚ź│²Ż¼ØŌČ╚Å─22.16 mg Īż L-1ĮĄĄ═ĄĮ3.80 mg Īż L-1;į┌9~14 hļAČ╬Ż¼Ž§╦ß¹}╚ź│²┌ģä▌£pŠÅŻ¼Ū·ŠĆą▒┬╩▀hąĪė┌6~9 hļAČ╬;ų▒ĄĮ17 hĢrŻ¼Ž§╦ß¹}╚ź│²┬╩▀_ĄĮ100%.┼cų«ī”æ¬Ą─╩ŪŻ¼į┌6~13 hļAČ╬Ż¼ü厧╦ß¹}ķ_╩╝│÷¼FĘe└█Ż¼ØŌČ╚Å─1.24 mg Īż L-1╔²Ė▀ų┴16.76 mg Īż L-1;ŠoĮėų°į┌13~14 hŻ¼ü厧╦ß¹}╚ź│²ų┴0.84 mg Īż L-1Ż¼╚ź│²┬╩ĮėĮ³100%.┴Ē═Ō╚ńłD 4b╦∙╩ŠŻ¼į┌Ž§╦ß¹}╚ź│²║═ü厧╦ß¹}Ęe└█▓ó▀MČ°┐ņ╦┘╚ź│²▀@ę╗ļAČ╬Ż¼ėąÖC╠╝Ž¹║─ĮėĮ³100%Ż¼▒Ē├„į┌║├č§Ę┤Ž§╗»Š·ū„ė├Ž┬Ż¼Ž§╦ß¹}ŅÉ╬’┘|║═ėąÖC╠╝▒╗Ž¹║─.Ą½╩Ūį┌Ž§╦ß¹}╚ź│²▀^│╠ųą░l╔·┴╦├„’@Ą─ü厧╦ß¹}Ęe└█Ż¼Ęe└█┴┐╩ŪŽ§╦ß¹}▀ĆįŁ┴┐Ą─72.80%Ż¼▀@┼cę╗ą®čąŠ┐░l¼FĄ─Ž§╦ß¹}╚ź│²▀^│╠ųą¤oü厧╦ß¹}Ęe└█Ą─ĮY╣¹ŽÓĘ┤.Ęų╬÷įŁę“╩ŪŻ¼ü厧╦ß▀ĆįŁ├Ė│ąō·īóü厧╦ß¹}▀ĆįŁĄ─╣”─▄Ż¼Ą½ü厧╦ß¹}į┌╚▄ę║ųąĢ■«a╔·ė╬ļxü厧╦ß(FNA)Ż¼FNAėą▌^ÅŖĄ─╔·╬’ČŠąįŻ¼ī”╬ó╔·╬’Ą─╔·ķL┤·ųx«a╔·ęųųŲū„ė├.▀@┐╔ęįĮŌßī×ķ╩▓├┤Ž§╦ß¹}Ą─┐ņ╦┘╚ź│²▀^│╠į┌9 h║¾▒╗ĮKų╣Ż¼9 h║¾╗∙▒Š╩ŪŠÅ┬²╚ź│²▀^│╠.┴Ē═ŌŻ¼║├č§Ę┤Ž§╗»Š·ę▓ąĶę¬║Ž│╔ČÓĘN├Ė║═╝Ü░¹ĮMĘųęį▀mæ¬ą┬│÷¼FĄ─ü厧╦ß¹}ŁhŠ│.«öŠ·╚║▀mæ¬ą┬Ą─Ą¬į┤║¾Ż¼āHį┌1 hā╚ü厧╦ß¹}╚ź│²┬╩╝┤ĮėĮ³100%.
ĪĪĪĪ3.2.2 Ę┤æ¬Ų„▀\ąąĀŅæB
ĪĪĪĪįć“×ųą░l¼FŻ¼š¹éĆĘ┤æ¬▀^│╠ųą¤oÖC╠╝ØŌČ╚├„’@╔²Ė▀Ż¼▀@ę▓įS┐╔ęįÅ─łD 4cųąšęĄĮįŁę“.łD 4c×ķĘ┤æ¬Ų„ā╚│ų└m18 hĄ─£žČ╚Īó╚▄ĮŌč§║═pHĄ─ūā╗»┌ģä▌.pH│╩ųØu╔Ž╔²┌ģä▌Ż¼Å─įć“×ķ_╩╝Ą─7.80╔²ų┴įć“×ĮY╩°Ą─8.53;Č°ļSų°pHųĄĄ─╔²Ė▀Ż¼╦«ųą╚▄ĮŌĄ─CO2į÷ČÓŻ¼¤oÖC╠╝ę▓ŽÓæ¬╔²Ė▀.┴Ē═ŌŻ¼×ķ┐╝▓ņĖ▀£žŚl╝■Ž┬║├č§Ę┤Ž§╗»Š·Ą─▀mæ¬─▄┴”Ż¼£žČ╚į┌įć“×╚½│╠ŠS│ų▌^Ė▀╦«ŲĮŻ¼═¼Ģrę▓╩╣CO2Ą─╚▄ĮŌąį╠ßĖ▀Ż¼▀Mę╗▓Į╠ßĖ▀┴╦╦«ųą¤oÖC╠╝Ą─ØŌČ╚Ż¼┼cłD 4bųąĄ─¤oÖC╠╝ØŌČ╚ūā╗»┌ģä▌Ę¹║Ž.╚▄ĮŌč§į┌6~14 hā╚▓©äė├„’@Ż¼┼c▀@ę╗ĢrķgČ╬Ž§╦ß¹}║═ü厧╦ß¹}Ą─┐ņ╦┘╚ź│²ę▓ŽÓ╬Ū║ŽŻ¼▒Ē├„║├č§Ę┤Ž§╗»Š·į┌╚ź│²Ž§╦ß¹}ŅÉ╬’┘|ĢrŻ¼ū„×ķļŖūė╩▄¾wĄ─O2ę▓═¼Ģr▒╗▀ĆįŁ.
ĪĪĪĪ3.3 ęįü厧╦ß¹}×ķĄ¬į┤ÅU╦«Ą─║├č§Ę┤Ž§╗»čąŠ┐
ĪĪĪĪ3.3.1 ╬’┘|╚ź│²▀^│╠┼cĘų╬÷ łD 5a║═5bĘųäe×ķĄ¬ŅÉ╬’┘|║═╠╝ŅÉ╬’┘|╚ź│²╝░▐D╗»┌ģä▌.ė╔łD 5a┐╔ų¬Ż¼į┌0~3 hļAČ╬Ż¼ü厧╦ß¹}ØŌČ╚ė╔20.92 mg Īż L-1ĮĄų┴13.26 mg Īż L-1Ż¼Ž§╦ß¹}ØŌČ╚ė╔9.20 mg Īż L-1╔²ų┴17.16 mg Īż L-1.▀@╩Ūė╔ė┌ü厧╦ß¹}ØŌČ╚▌^Ė▀ĢršTī¦┴╦ü厧╦ßč§╗»├ĖŻ¼īó▓┐Ęųü厧╦ß¹}č§╗»×ķŽ§╦ß¹}(Åł┼Óė±Ą╚Ż¼2010);ü厧╦ß¹}▒╗č§╗»Ą─┴┐▒╚Ž§╦ß¹}Ą─į÷╝ė┴┐╔┘0.3 mg Īż L-1Ż¼▒Ē├„ėąśO╔┘┴┐Ą─ü厧╦ß¹}▒╗║├č§Ę┤Ž§╗»Š·▀ĆįŁŻ¼▀@┼cłD 5bųą▀@ę╗ĢrķgČ╬ėąÖC╠╝Ą─╔┘┴┐Ž¹║─ŽÓĘ¹.
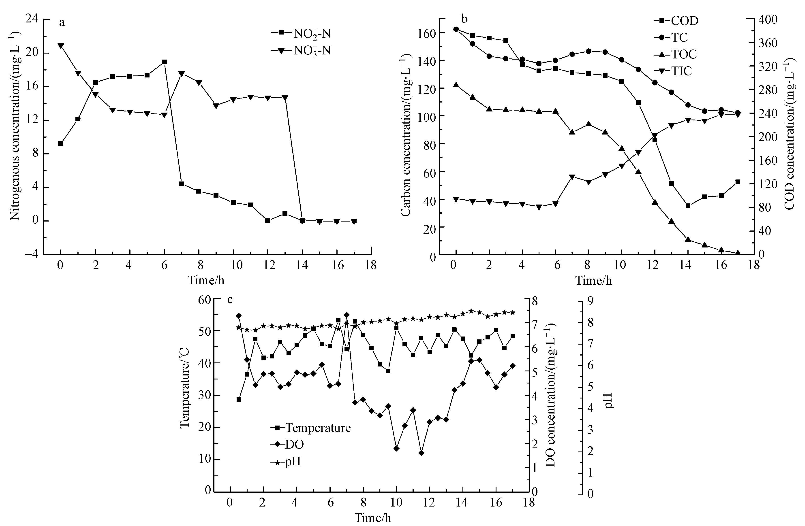
ĪĪĪĪłD5 ╬’┘|ØŌČ╚║═▀\ąąģóöĄūā╗»┌ģä▌
ĪĪĪĪį┌3~6 hļAČ╬Ż¼ü厧╦ß¹}ØŌČ╚╬óŽ┬ĮĄŻ¼Ž§╦ß¹}ØŌČ╚ąĪĘ∙╔Ž╔².6~7 hā╚Ž§╦ß¹}ØŌČ╚┤¾Ę∙Č╚Ž┬ĮĄŻ¼╚ź│²┬╩▀_ĄĮ80%ęį╔ŽŻ¼═¼Ģrü厧╦ß¹}ØŌČ╚╔²ų┴Ė▀³c.Ęų╬÷įŁę“Ż¼ę╗╩Ūė╔ė┌Ž§╦ß¹}Š▀ėą▌^Ė▀Ą─č§╗»▀ĆįŁļŖ╬╗Ż¼└¹ė├Ųõū„×ķļŖūė╩▄¾wĢr╗∙┘|ßīĘ┼Ą──▄┴┐▌^Ė▀;┴Ē═ŌFretteĄ╚šJ×ķ└Ēšō╔ŽŽ§╦ß¹}▀ĆįŁ«a╔·Ą──▄┴┐╩Ūü厧╦ß¹}▀ĆįŁ«a╔·─▄┴┐Ą─3.8▒ČŻ¼Ė∙ō■ā׎╚└¹ė├įŁätŻ¼«ö╦«ųą═¼Ģr┤µį┌Ž§╦ß¹}║═ü厧╦ß¹}ĢrŻ¼Ę┤Ž§╗»Š·ā׎╚└¹ė├Ž§╦ß¹}▀Mąą║├č§Ę┤Ž§╗».Č■╩Ūė╔ė┌ü厧╦ß¹}╩ŪŽ§╦ß¹}Ę┤Ž§╗»▀^│╠Ą─ųąķg«a╬’Ż¼ŲõØŌČ╚▀^Ė▀Ģręūī”Ž§╦ß¹}▀ĆįŁ×ķü厧╦ß¹}▀@ę╗▓Į¾E«a╔·ęųųŲū„ė├Ż¼Å─Č°ĮĄĄ═Ž§╦ß¹}Ą─▀ĆįŁ┴┐Ż¼▀@┐╔ęįĮŌßī×ķ╩▓├┤Ž§╦ß¹}┐ņ╦┘╚ź│²ų╗│ų└m┴╦1 hŠ═▀M╚ļŠÅ┬²╚ź│²▀^│╠Ż¼Č°▀@éĆĢrķg³cš²╩Ūü厧╦ß¹}ØŌČ╚Ą─Ė▀³c.šf├„▀^Ė▀Ą─ü厧╦ß¹}ØŌČ╚ęųųŲ┴╦Ž§╦ß¹}Ą─▀ĆįŁ▀^│╠.ļS║¾Ą─7~14 hā╚Ż¼Ž§╦ß¹}╚ź│²▀_ĄĮ100%;ļSų°Ž§╦ß¹}Ą─╚ź│²ĮėĮ³═Ļ│╔Ż¼ü厧╦ß¹}╚ź│²Å─13 hķ_╩╝Ż¼1 hā╚╚ź│²┬╩╝┤▀_ĄĮ100%.╚ńŪ░╦∙╩÷Ż¼ü厧╦ß¹}«a╔·Ą─FNAėąČŠŻ¼║├č§Ę┤Ž§╗»Š·ąĶę¬Ģrķg║Ž│╔├Ė║═╝Ü░¹ĮMĘųŻ¼▀@ę╗▀^│╠│ų└m┴╦11 h.Č°Ūęį┌═¼Ģr┤µį┌Ž§╦ß¹}║═ü厧╦ß¹}Ą─ŁhŠ│ųąŻ¼Č■š▀╚ź│²Ą─╦┘Č╚(┐é╣▓14 h)▒╚å╬ę╗Ž§╦ß¹}(18 h)ĢrĖ³┐ņ.
ĪĪĪĪį┌Ę┤æ¬▀^│╠ųąŻ¼▓┐ĘųĢrķg³cÖz£yĄĮ╬ó┴┐Ą─░▒Ą¬┤µį┌.ļm╚╗╔┘┴┐Ą─░▒Ą¬Ģ■┤┘▀M▀ĆįŁ├ĖĄ─║Ž│╔║═Ž§╦ß¹}ŅÉĄ─╚ź│²Ż¼Ą½░▒Ą¬ų╗į┌▓┐ĘųĢrķg³cÖz£yĄĮŻ¼¤o╠žČ©Ą─ęÄ┬╔Ż¼╦∙ęį▀@ĘN┤┘▀Mū„ė├╩Ūʱ┤µį┌ė┌▒Šįć“×蹊┐ųą▓ó▓╗─▄┤_Č©.
ĪĪĪĪ3.3.2 Ę┤æ¬Ų„▀\ąąĀŅæB
ĪĪĪĪłD 5c×ķĘ┤æ¬Ų„ā╚│ų└m24 hĄ─£žČ╚Īó╚▄ĮŌč§║═pHĄ─ūā╗»┌ģä▌.įć“×ųąŻ¼pH│╩ųØu╔Ž╔²┌ģä▌Ż¼Å─įć“×ķ_╩╝Ą─7.71╔²ų┴įć“×ĮY╩°Ą─8.41Ż¼▒Ē├„Ę┤Ž§╗»ū„ė├į┌│ų└mĘĆČ©▀Mąą.╚▄ĮŌč§į┌6~14 hā╚▓©äė├„’@Ż¼┼c▀@ę╗ĢrķgČ╬Ž§╦ß¹}║═ü厧╦ß¹}Ą─┐ņ╦┘╚ź│²ŽÓ╬Ū║Ž.▒Ē├„║├č§Ę┤Ž§╗»Š·į┌╚ź│²Ž§╦ß¹}ŅÉ╬’┘|ĢrŻ¼ū„×ķļŖūė╩▄¾wĄ─O2ę▓═¼Ģr▒╗▀ĆįŁŻ¼▀@┼cŪ░╬─Ž§╦ß¹}×ķĄ¬į┤Ą─įć“×¼FŽ¾ę╗ų┬.
ĪĪĪĪ3.4 ║├č§Ę┤Ž§╗»Š·Ą─ĶbČ©
ĪĪĪĪ╬█─ÓśėŲĘ╚Īūį║├č§Ę┤Ž§╗»įć“×ĮY╩°║¾Ą─┴„ļx╠Ņ┴Ž╔·╬’─ż.łD 6×ķæ¬ė├Ę┤Ž§╗»Š·╗∙ę“Ų¼Č╬║═GenBanköĄō■Äņ╦∙½@Ą├Ą─Ę┤Ž§╗»Š·ŽĄĮy░lė²śõ. Öz£yĄĮĄ─Ę┤Ž§╗»Š·Įø╬─½I▒╚ī”Ą├ų¬Ż║ŲõųąFN555565.1Īó FN555559.1ĪóFN555558.1Ż¼AY078272.1ĪóAY078256.1ĪóAM230913.1ĪóAM230896.1×ķ║├č§Ę┤Ž§╗»Š·;GQ384052.1ĪóEF558380.1ĪóEF558490.1×ķ«ÉBŽ§╗»-║├č§Ę┤Ž§╗»Š·;Ųõ╦³╝ÜŠ·Ģ║Ģr×ķąįĀŅ▓╗├„Ą─Ę┤Ž§╗»Š·Ż¼Ģ║Ģr¤o╬─½I▒╚ī”ĮY╣¹.ĶbČ©ĮY╣¹▒Ē├„Ż¼ļSų°╦«ŁhŠ│ųąĄ¬į┤ŅÉą═Ą─ūā╗»Ż¼┴„ļx╠Ņ┴Ž╔·╬’─żā╚║├č§Ę┤Ž§╗»Š·Ą─ĘNī┘░l╔·ūā╗»ęį▀mæ¬ą┬Ą¬į┤ŁhŠ│Ż¼┼c▒Šįć“×蹊┐ķ_╩╝Ģr╦∙ė├Ą─┴„ļx╠Ņ┴Ž╔·╬’─żā╚Ą─Š·ī┘ėąę╗Č©ģ^äe;▓┐ĘųŠ·╚║Š▀éõ«ÉBŽ§╗»─▄┴”Ż¼┐╔ęįĮŌßīį┌╠Ä└ĒĄ═╠╝Ą¬▒╚ÅU╦«Ģr0~10 hā╚┐饬║═ėąÖC╠╝į┤ūā╗»┌ģä▌Ą─ŽÓĻPąį.Š▀¾wģóęŖ╬█╦«īÜ╔╠│Ū┘Y┴Ž╗“http://www.dongaorq.cnĖ³ČÓŽÓĻP╝╝ąg╬─ÖnĪŻ
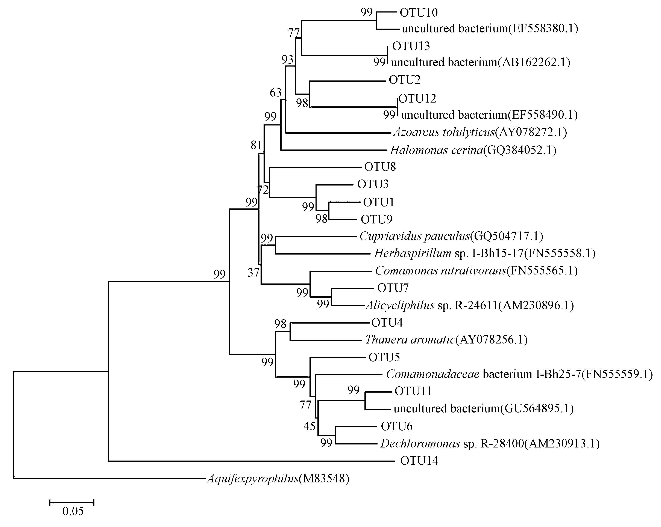
ĪĪĪĪłD6 Ę┤Ž§╗»Š·Ą─ŽĄĮy░lė²▀M╗»śõ
ĪĪĪĪ4 ĮYšō
ĪĪĪĪ蹊┐┴╦╔·╬’─żā╚╣”─▄Š·╚║×ķš¹¾w蹊┐║├č§Ę┤Ž§╗»╠žš„.į┌Ą═╠╝Ą¬▒╚ÅU╦«Ą─įć“×蹊┐ųą░l¼FŻ¼Ę┤æ¬ļAČ╬▒╗Ęų×ķ«ÉBŽ§╗»-║├č§Ę┤Ž§╗»║═ūįBŽ§╗»-║├č§Ę┤Ž§╗»ā╔ļAČ╬Ż¼Ą½Š∙Ę¹║Ž═¼▓ĮŽ§╗»Ę┤Ž§╗»╠žš„.į┌ęįŽ§╦ß¹}║═ü厧╦ß¹}Ęųäe×ķĄ¬į┤Ą─ÅU╦«Ę┤Ž§╗»įć“×蹊┐░l¼FŻ¼║├č§Ę┤Ž§╗»Š·ī”Ą¬į┤Ą─└¹ė├ėąŽ╚║¾ų«Ęų;Ž§╦ß¹}╚ź│²▀^│╠ųąĢ■│÷¼Fü厧╦ß¹}Ą─Ęe└█Ż¼▀@╩Ū▓╗═¼▀ĆįŁ├Ėų«ķgŽÓ╗źęųųŲĄ─ū„ė├.į┌ü厧╦ß¹}║═Ž§╦ß¹}═¼Ģr┤µį┌Śl╝■Ž┬Ż¼Č■š▀╚ź│²╦┘┬╩(14 h)▒╚å╬ę╗Ž§╦ß¹}(18 h)Ą─╚ź│²Ė³┐ņ;║├č§Ę┤Ž§╗»Š·ī”Ą¬į┤Ą─Ė─ūāėąę╗éĆ▀mæ¬▀^│╠.ęį╔ŽĮYšōŠ∙į┌Ė▀£ž(>40 Īµ)Śl╝■Ž┬Ą├│÷.ŠC╔ŽŻ¼ęįš¹¾wŠ·╚║┤·╠µå╬ę╗Š·ųĻĄ─║├č§Ę┤Ž§╗»įć“×蹊┐Ż¼▓óį┌ŽÓī”É║┴ėĄ─╦«ŁhŠ│Ž┬▀MąąŻ¼ėąų·ė┌┐╝▓ņ║├č§Ę┤Ž§╗»Š·æ¬ė├ė┌īŹļH╬█╦«╠Ä└ĒįO╩®ųąĄ─┐╔─▄ąį.

